
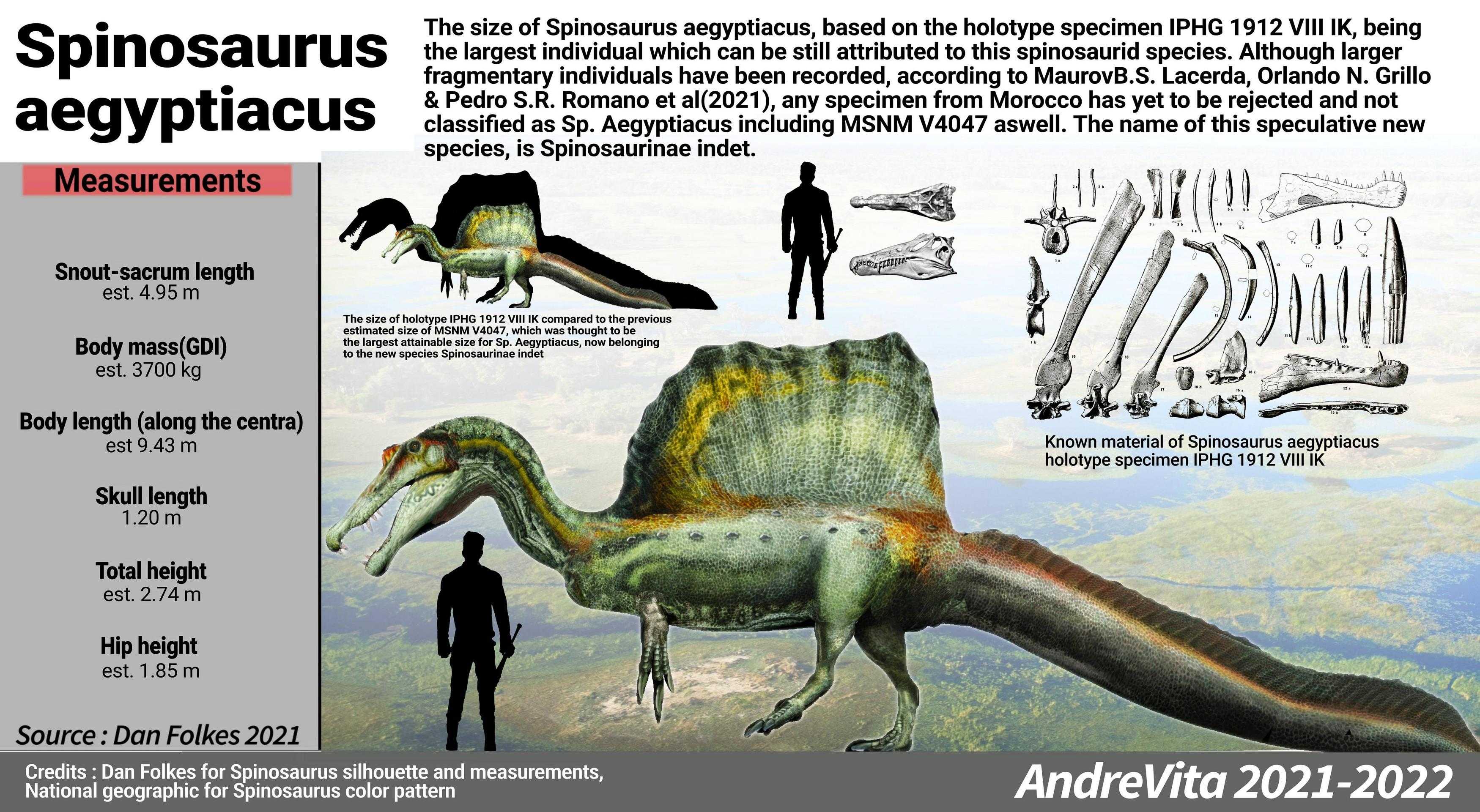
Древний мир
Разные виды динозавров жили на Земле в разное время: одни виды вымирали, появлялись другие. В целом эра этих существ продлилась более 150 миллионов лет. Если сравнить с этим периодом время существования человека, то мы живем всего 200 000 лет. По официальной версии, люди и динозавры жили в разные эры, но вот наскальные рисунки, различные археологические находки, ставят под сомнение эту версию.
Само слово «динозавр» обозначает ужасная или страшная ящерица. Эти существа были хладнокровными, нуждались в большом количестве солнечного света. В официальной версии того, как появились динозавры, говорится, что эти существа эволюционировали из рептилий, которые жили на планете до них. Предки динозавров жили и на суше, и в воде. Их называют архозавры – древние ящерицы. Они выглядели как амфибии, питались растительной пищей. У древних ящеров были легкие, они откладывали яйца на суше. Постепенно рептилии становились крупнее, хотя первые динозавры были мелкими, размером с курицу. Некоторые виды такими оставались на протяжении всего своего существования. Другие же виды стали крупными, они увеличивались в размерах, крепли. Так появились динозавры, которые изменялись и совершенствовались на протяжении миллионов лет, заселяя Землю.
Classification
Most analyses place Afrovenator within the Megalosauridae, which was formerly a «wastebasket family» which contained many large and hard-to-classify theropods, but has since been redefined in a meaningful way, as a sister taxon to the family Spinosauridae within the Megalosauroidea.
A 2002 analysis, focused mainly on the noasaurids, found Afrovenator to be a basal megalosaurid. However, it did not include Dubreuillosaurus (formerly Poekilopleuron valesdunensis), which could affect the results in that region of the cladogram (Carrano et al. 2002).
Other more recent and more complete cladistic analyses show Afrovenator in a group of Megalosauridae with Eustreptospondylus and Dubreuillosaurus. This group is either called Megalosaurinae (Allain 2002) or Eustreptospondylinae (Holtz et al. 2004). The latter study also includes Piatnitzkysaurus in this taxon. A study by Matthew Carrano from 2012 placed Afrovenator in a megalosaurid Afrovenatorinae.
A few alternative hypotheses have been presented for Afrovenators relationships. In Sereno’s original description, Afrovenator was found to be a basal spinosauroid (he at the time used the name «Torvosauroidea»), outside of Spinosauridae and Megalosauridae (which he called «Torvosauridae»). Finally, another recent study places Afrovenator outside of the Megalosauroidea completely, and instead finds it more closely related to Allosaurus (Rauhut 2003). This is the only study to draw this conclusion.
In a revision of Carnosauria lead by the discovery of the basal Allosauroid Asfaltovenator recovering a paraphyletic Megalosauroidea, but was latter re-recovered as a monophyletic group, with Afrovenator alone, and taxa referred previously to Afrovenatorinae such as Dubreuillosaurus, Magnosaurus and Piveteausaurus clading with Eustreptospondylus.
Discovery and naming
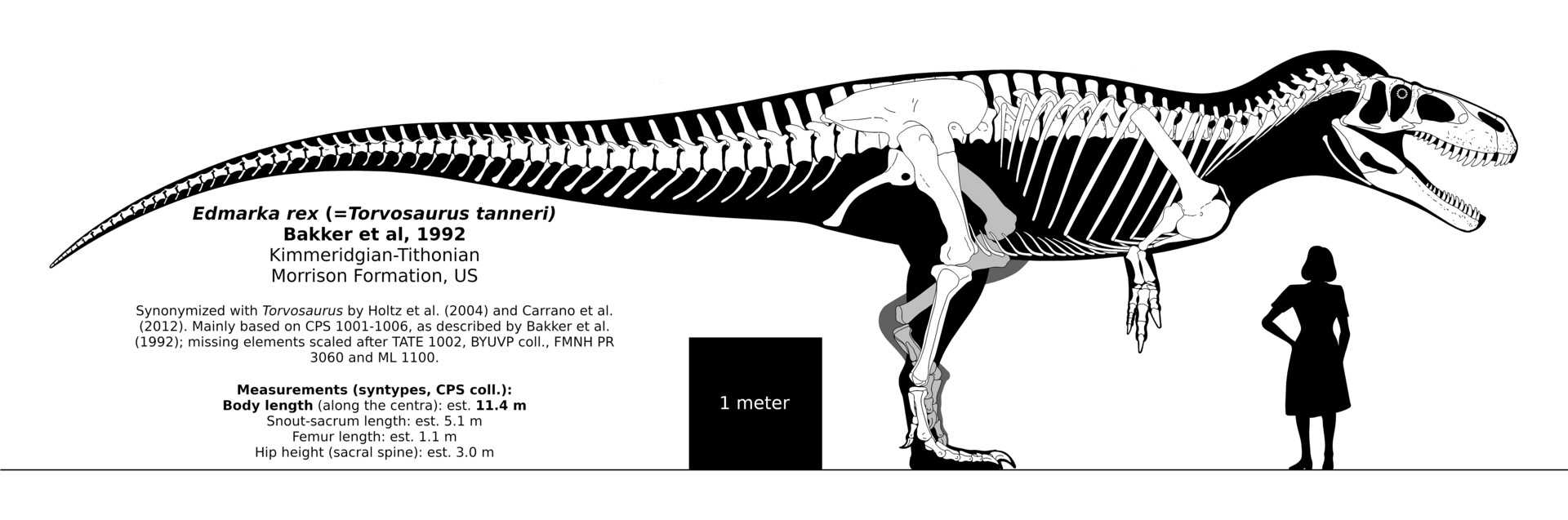
Skeletal diagram of known material
The remains of Afrovenator were discovered in 1993 in the Tiourarén Formation of the department of Agadez in Niger. The Tiourarén was originally thought to represent the Hauterivian to Barremian stages of the early Cretaceous Period, or approximately 132 to 125 million years ago (Sereno et al. 1994). However, re-interpretation of the sediments showed that they are probably Middle to Late Jurassic in age, dating Afrovenator to the Bathonian to Oxfordian stages, between 167 and 161 mya. The sauropod Jobaria, whose remains were first mentioned in the same paper which named Afrovenator, is also known from this formation.
Afrovenator is known from a single relatively complete skeleton, holotype UC OBA 1, featuring most of the skull minus its top (likewise the mandible, or lower jaws, are lacking apart from the prearticular bone), parts of the spinal column, partial forelimbs, a partial pelvis, and most of the hind limbs. This skeleton is housed at the University of Chicago.
The generic name comes from the Latin afer, «African», and venator, «hunter». There is one named species, Afrovenator abakensis. The generic name refers to its predatory nature, and its provenance from Africa. The specific name refers to Abaka, the Tuareg name for the region of Niger where the fossil was found. The original short description of both genus and species is found in a 1994 paper which appeared in the prestigious journal Science. The primary author was well-known American paleontologist Paul Sereno, with Jeffrey Wilson, Hans Larsson, Didier Dutheil, and Hans-Dieter Sues as coauthors.
Recent discoveries in the region include referred teeth (MUPE HB-118, 125, 142) from the underliying Irhazer II Formation and TP4-12, a rostral part of left maxilla from the Tiourarén Formation at NE Tadibene.
The Dinosaur in Detail
The Afrovenator had several unique features, such as its long humerus and lightweight build, that suggest a dinosaur that was built for speed. These would have been influential adaptations for a carnivorous dinosaur, allowing it to chase down prey in its environment.
One of the most notable specimens is the nearly complete skeleton found in Niger. This specimen has provided invaluable insights into the Afrovenator’s anatomy and lifestyle. For example, the skeleton shows that the Afrovenator had a particularly long humerus, or upper arm bone, which would have given it a greater reach. This could have been a crucial advantage when hunting prey.
The skull was rather flat with a small maxillary fenestra and a distinctive rounded horn on top of the lacrimal bone near the eyes. The lower branch of the postorbital bone behind the eyes was transversely wide and the jugal bone near the cheek was short, deep, and pneumatized–meaning it was filled with air cavities. These features suggest a dinosaur with a keen sense of sight and possibly a complex social or mating behavior.
Наука о динозаврах
Изучение динозавров — часть научной дисциплины, которая называется «палеонтология» . Палеонтологи анализируют ископаемые останки растений и животных, чтобы лучше понимать эволюцию жизни на Земле и взаимодействие живых организмов друг с другом. Наука изучает два подтипа живых существ: позвоночные и беспозвоночные. Исследователи первого вида изучают ископаемых животных, которые обладали позвоночником или хордой (костным или хрящевым хребтом, поддерживающим тело). К ним относятся динозавры, рыбы, ящерицы, змеи, птицы. К беспозвоночным, которых изучают палеонтологи, относятся улитки, трилобиты, устрицы, моллюски, кальмары.
Меловой период
Меловой период — это период, в котором динозавры достигли пика разнообразия. И птицетазовые и ящеротазовые обзавелись огромным количеством новых видов. Это был самый длинный период мезозоя, в течение которого Земля начала принимать современный вид. На протяжении мелового периода на суше, в море и в воздухе доминировали рептилии, млекопитающие в это время занимали скромное положение.
География и климат мелового периода
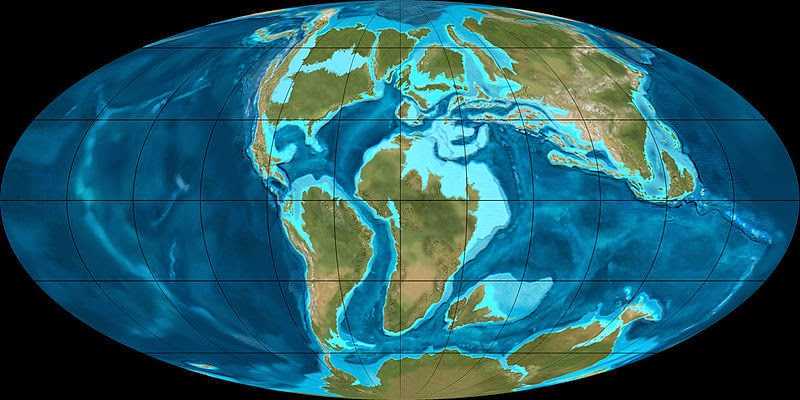 Поверхность Земли в меловой период
Поверхность Земли в меловой период
В начале мелового периода Пангея продолжала распадаться на части. Из ее осколков формировались Северная и Южная Америки, Европа, Азия и Африка. Центральная часть Северной Америки была занята Западным внутренним морем (которое дало огромное количество окаменелостей морских рептилий). Индия была гигантским островом в океане Тетис. Климат, в основном, был таким же жарким и влажным, как и в юрский период. В меловом периоде наблюдалось повышение уровня моря и заболачивание обширных территорий.
Наземная жизнь в меловой период
Динозавры достигли своего рассвета во время мелового периода. В течение 80 миллионов лет тысячи видов обитали на медленно разделяющихся континентах. Среди них были хищники такие как, тиранозавры и другие разновидности теропод, в том числе быстроногие орнитомимиды, пернатые теризинозавры и огромное количество мелких пернатых динозавров. Большинство травоядных зауроподов обитавших в юрском периоде вымерло, однако они дали начало новым разновидностям динозавров. Титанозавры распространились по всем континентам кроме Атарктиды и стали еще крупнее. Огромные стада цератопсов и гадрозавры распространились на равнинах Северной Америки и Евразии. Помимо них, среди травоядных были анкилозавры и пахицефалозавры.

На протяжении мезозойской эры млекопитающие не могли конкурировать с динозаврами. Это были мелкие животные обитающие в кронах деревьев или в норах под землей. Тем не менее стали появляться и более крупные животные. Например был десяти-килограммовый репеномам (Repenomamus), который охотился на мелких динозавров.
Морская жизнь в меловой период
В раннем меловом периоде исчезли ихтиозавры. На их месте появились мозазавры, огромные плиозавры (кронозавр), и несколько меньшие по размерам плезиозавры (эласмозавр). Костисты рыбы, появившиеся в триасе, в меловом периоде становятся многочисленны и широко распространяются в Мировом Океане. В морях также встречалось много акул. После исчезновения морских рептилий, доминирующее положение в океанах займут рыбы.
Летающие животные в меловой период
К концу мелового периода птерозавры достигли огромных размеров. Например кетцалькоатль с размахом крыльев в 10 метров. Это был период угасания птерозавров, их постепенно вытесняли первые настоящие птицы. Эти ранние птицы произошли от наземных пернатых динозавров, а не птерозавров, и были лучше приспособлены к изменяющимся климатическим условиям.
Жизнь растений в меловой период
Что касается растений, то наиболее важным эволюционным событием мелового периода было быстрое видообразование цветковых растений. Земля была покрыта густыми лесами. Растительность не только обеспечивала динозавров пищей, но и способствовала коэволюции большого количества насекомых, особенно жуков.
Виды динозавров
Все известные виды динозавров делятся на два больших отряда: птицетазовые и ящеротазовые. Эти отряды имеют свои классификации. Так, выделяют группы водных, летающих, травоядных, плотоядных динозавров и некоторые другие. А откуда взялись динозавры и почему их разделили на группы?
С тех пор как ученые стали находить останки водных представителей, возник вопрос, а как рождаются динозавры этой группы? Те, которые передвигаются по суше, летают, обычно откладывают яйца, а размножение водных происходит или путем выхода существа на сушу для откладывания яиц, как это делают современные черепахи, или же были иные методы размножения.
Среди водных обитателей, самыми необычными являются:
- Плиозавр. Он считается хищником, питался всем, что видел. У этого существа не было врагов, из-за чего он первым нападал.
- Ихтиозавр. Первые останки этих существ были найдены на территории России, но больше всего было находок в Германии. Эти ящеры жили стаями, поэтому могли с легкостью дать отпор врагам, и им легче было добывать пищу.
- Мозазавр. Останки этих завров были найдены по всей планете, даже в холодной Антарктиде. Считается, что от этого вида произошли вараны. Мозазавр жил в воде, передвигался по-змеиному, как морские угри.
- Эласмозавр. Он считается необычным хищником, который мог охотиться с любого расстояния. Этот динозавр без особого труда ловил любую добычу, даже самую быструю.
- Шонизавр. Среди водных рептилий это существо считается самым крупным. Оно питалось кальмарами, моллюсками, некоторыми видами осьминогов.
Это далеко не все представители водного мира, которые населяли планету миллионы лет назад.
К летающим видам относятся птерозавр, археоптерикс, птеродактиль, орнитохейрус. Некоторые из них достигали размера маленького самолета.







Самыми популярными хищными динозаврами являются тиранозавр, мегалозавр, спинозавр, тарбозавр, гиганотозавр, велоцираптор.
Представителями травоядных являются стегозавр, брахиозавр, диплодок, бронтозавр, трицератопс, игуанодон.
Группы динозавров
Зауроподы — группа растительноядных динозавров, которая отличалась самыми внушительными размерами среди остальных — крупным телом, длинным хвостом и шеей, которая, как и жирафу, помогала доставать древесную листву для питания.
Тероподы — плотоядные динозавры. Эта группа хищников обладала великолепным зрением, острыми зубами и не менее острыми когтями, которые очень помогали при охоте.
Крупные травоядные динозавры с огромными костяными пластинами, расположенными вдоль длинного позвоночника. Считается, что эти шипы служили не только устрашением для хищников, визуально увеличивая размеры динозавра, но и участвовали в процессах терморегуляции благодаря огромному количеству расположенных в них кровеносных сосудов.
Брахиозавры — огромные динозавры, которые жили стадами. Их размеры можно сопоставить с двумя двухэтажными автобусами, поставленными друг на друга. Как и все зауроподы, они обладали длинной шеей, помогающей им доставать сочную листву с высоких деревьев.
Часть динозавров предпочитала передвигаться исключительно на двух ногах, поэтому их назвали двуногими, другие же ходили только на четырех. Но встречались виды, которые свободно могли передвигаться как на двух, так и на четырех ногах.
Further reading
- Allain, R (2002). «Discovery of megalosaur (Dinosauria, Theropoda) in the middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae». Journal of Vertebrate Paleontology 22 (3): 548–563. doi:10.1671/0272-4634(2002)022.
- Carrano, M.T.; Sampson, S.D.; Forster, C.F. (2002). «The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar». Journal of Vertebrate Paleontology 22 (3): 510–534. doi:10.1671/0272-4634(2002)022.
- Holtz, T.R., Molnar, R.E., Currie, P.J. (2004). «Basal Tetanurae». In: Weishampel, D.B., Dodson, P., & Osmolska, H. (eds.). The Dinosauria (2nd edition). Berkeley: University of California Press. pp. 71–110.
- Rauhut, O.W.M. (2003). The Interrelationships and Evolution of Basal Theropod Dinosaurs. Special Papers in Palaeontology 69. London: The Palaeontological Association. pp. 1–215.
Wikidata ☰ Q131155 entry
Библиография
- Аллен, Р. (2002). «Открытие мегалозавра (Dinosauria, Theropoda) в среднем бате Нормандии ( Франция ) и его значение для филогении базальных столбняков». Журнал палеонтологии позвоночных, 22 (3): 548-563.
- Бенсон, Р.Б.Дж., Барретт, П.М., Пауэлл, Х.П., и Норман, Д.Б. (2008). «Таксономический статус Megalosaurus bucklandii (Dinosauria, Theropoda) из средней юры Оксфордшира, Великобритания». Палеонтология, 51 (2): 419-424.
- Бенсон, RBJ (2010). «Описание Megalosaurus bucklandii (Dinosauria: Theropoda) из бата Великобритании и взаимоотношений теропод средней юры». Зоологический журнал Линнеевского общества . doi: 10.1111 / j.1096-3642.2009.00569.x.
- Бенсон, Р.Б.Дж., Каррано, М.Т. и Брусатте, С.Л. (2010). «Новая клада архаичных крупных хищных динозавров (Theropoda: Allosauroidea), дожившая до позднего мезозоя». Naturwissenschaften 97 (1): 71–78.
The Afrovenator in its Natural Habitat and Environment
This dinosaur lived during the Middle Jurassic period, a time of significant evolutionary diversification among dinosaurs. The African environment it inhabited was likely diverse and dynamic, offering a range of opportunities and challenges for this dinosaur.
As a carnivore, the Afrovenator would have been at the top of the food chain in its environment. Its diet likely consisted of smaller dinosaurs and possibly other animals. Given its body type, agility and speed would have made it a successful predator that could outrun its prey.
Its social behavior is less clear, but given its size and the nature of its environment, it’s possible that it was a solitary hunter. However, this is purely speculative, and more research is needed to confirm this.
Systematics
The result of the cladistic analysis carried out in the context of the first description (Sereno et al., 1994) shows Afrovenator as a representative of the extensive theropod clade Tetanurae . Within this clade, it appears as the most basal representative of the most basal Tetanurae clade «Torvosauroidea» (Spinosauroidea, Megalosauroidea) , which, in addition to Afrovenator, also includes the Spinosauridae and Torvosauridae (Megalosauridae) , the two last-named taxa being differentiated from Afrovenator by the presence of a characteristic sickle-shaped distinguish the last (distal) phalanx of the first finger of the hand. Tetanurae synapomorphies of the Afrovenator skull include in this analysis the presence of a maxillary fossa and the position of the rearmost tooth position in front of the front edge of the eye socket, Tetanurae synapomorphies of the postcranial skeleton are the «semilunate» carpale, the (presumably) atrophied third finger, the Wing-like or blade-like anterior trochanter of the femur and the lower ascending process of the astragalus (see ).
The results of the following analyzes mostly confirm the position of Afrovenator as a basal tetanur or spinosauroid / megalosauroid, albeit with variable relationships to the Spinosauridae and Megalosauridae. In addition, due to the modification and increasing size of the data sets (both with regard to the number of characteristics and with regard to the number of included taxa) and also due to the revision of the characteristic partly different synapomorphies for the tetanurae and their subclades than those in mentioned in the first description.
Position of Afrovenator within the “higher” theropods (greatly simplified), according to Sereno et al. (1994)
|
Position of Afrovenator within the Megalosauroidea (Spinosauroidea), according to Benson (2010)
|
Position of Afrovenator within the Megalosauroidea (Spinosauroidea), after Carrano et al. (2012)
|
For example, Benson (2010) and Carrano et al. (2012) the Jugale of Afrovenator in their data matrices contrary to the information in Sereno et al. (1994) as unpneumatized (features 34 and 52 respectively with feature status 0) and also for the wing-shaped anterior trochanter the coding (Benson, 2010: feature 191, condition 0; Carrano et al. (2012: feature 308, condition 0)) does not correspond the information in Sereno et al. (1994), although Benson (2010) explicitly states that he obtained the data on Afrovenator from its first description. In addition, Benson (2010) explicitly states that Megalosaurus is the only genus within the Megalosauridae, who have a pneumatized Jugale. A pneumatized Jugale is therefore not listed here as a feature of Afrovenator .
For example, the presence of a large maxillary fossa in Benson (2010: characteristic 15, state 1) is a synapomorphism of the common clade of Ceratosauria and Tetanurae (= Neotheropoda ; 0 → 1) and the presence of a «real» maxillary window (characteristic 15, state 2) a synapomorphism of the Neotetanurae (1 → 2; in each case only under the ACCTRAN premise) or the somewhat less inclusive common clade of Allosauroidea + Coelurosauria (0 → 2; only under the DELTRAN premise). In Carrano et al. (2012) is, however, a «real» Maxillarfenster a synapomorphy the Tetanurae while at the Megalosauroidea (incl. Afrovenator (Engl.) As a relatively basal clade of Tetanurae a reversal reversal of this state) is carried through to Maxillarfossa. The presence of an enlarged terminal phalanx of the first finger of the hand is in Benson (2010: characteristic 159, state 1) a synapomorphism of his «New clade B» (mainly Spinosauridae + Megalosauridae), but in this hypothesis Afrovenator is within this clade (and although within the Megalosauridae) in a sister group relationship with Dubreuillosaurus , which is characterized by a reversal with regard to this feature (i.e., unlike in Sereno et al., 1994, secondary loss within the Megalosauridae instead of a plesiomorphic state within the Megalosauroidea, although only under ACCTRAN- Premise). In Carrano et al. (2012) it is similar. In his case, the ingroup within the Megalosauridae, identified by the reversal (characteristic 259, state 0, only under ACCTRAN premise), to which Afrovenator belongs and which he calls Afrovenatorinae, comprises more than just two terminal taxa.
Material and find location
The known fossil material from Afrovenator has so far been limited to the holotype of the type species (collection number: MNN TIG1 formerly UC OBA 1), a relatively complete skeleton, which includes a partial upper skull (cranium) without premaxillary, frontal, parietal and square jugals as well as a pre-articular (non-tooth bearing lower jaw bone), both humerus bones, both almost complete hands, almost complete pelvis, legs and parts of the cervical, trunk and tail spine.
These remains were discovered in the Tamerát fossil locale, about 100 km west-southwest of Agadez in the Tiouarén Formation of the Iullemmeden Basin in Niger and were first scientifically described in 1994 by Paul Sereno , Jeffrey Wilson, Hans Larsson, Didier Dutheil and Hans-Dieter Sues .
The sediments of the find layer represent an alluvial plain in a semi-humid or humid climate. Traditionally regarded as Early Cretaceous , which made Afrovenator abakensis the first relatively complete theropod from the Cretaceous Africa at the time of the first description, the Tiouarén Formation was based on the dinosaur fauna it contained in 2009 to a late Middle to early Upper Jurassic age ( Bathon — Callov ,? Oxford ) newly dated.
Описание
Размер афровенатора (оранжевого цвета) по сравнению с двумя другими афровенаторами
Судя по одному известному скелету, этот динозавр был около 8 метров (26 футов) в длину, от морды до кончика хвоста, и, по словам Грегори С. Пола , имел вес около 1 тонны . Томас Р. Хольц-младший оценил его в 7,6 метра (25 футов) в длину и в 453–907 кг (1 000–2 000 фунтов) в весе. В 2016 году он получил более низкую оценку: 6,8 метра (22,3 фута) в длину, 1,9 метра (6,2 фута) в высоту по бедрам и 790 кг (1,742 фунта) в весе. Серено подчеркнул, что общее телосложение было грациозным и что передние конечности и голень были относительно длинными: длина плечевой кости составляла сорок сантиметров, а голени и четвертой плюсневой кости — 687 и 321 миллиметр соответственно, по сравнению с длиной бедренной кости семьдесят шесть. сантиметры.
Было установлено несколько аутапоморфий , которые отличают Афровенатор от его ближайших родственников. Впадина, в которой находится анторбитальное отверстие , имеет передний конец в виде лопасти. Третий шейный позвонок имеет невысокий прямоугольный позвоночник. Кость запястья в форме полумесяца очень плоская. Первая пястная кость имеет широко расширяющуюся поверхность контакта со второй пястной костью. Ножка лобковой кости сзади насечена.
В целом череп довольно плоский, его высота меньше трехкратной длины, что не может быть точно определено из-за отсутствия передних челюстей . На верхней челюсти , имеющей длинную переднюю ветвь, имеется четырнадцать зубов, как можно заключить по лункам: сами зубы были потеряны. Имеется небольшое верхнечелюстное отверстие, не доходящее до края анторбитальной впадины и расположенное за переднечелюстным отверстием. Слезная кость имеет характерный закругленный рог на вершине. Нижняя ветвь заглазничной кости поперечно широкая. Скуловая кость короткая, глубокая и pneumatised.
Триасовый период
250 миллионов лет назад, в начале Триасового периода, жизнь на Земле восстанавливалась после пермско-триасового вымирания, в ходе которого исчезло более 75% наземных и 95% морских видов. Значимыми событиями развития животной жизни в Триасе были выделение из архозавров птерозавров, крокодилов и ранних динозавров, а также эволюция терапсид в первых настоящих млекопитающих.
Климат и география Триасового периода
Поверхность Земли в триасовый период
На протяжении триасового периода все континенты Земли были объединены в обширный супер-континент, называемый Пангея (окруженный огромным океаном Панталасса). Ледники на полюсах отсутствовали, в экваториальной зоне климат был жарким и сухим, с сильными муссонами. По некоторым оценкам, на большей части Пангеи средняя температура воздуха превышала 37,7 градусов по Цельсию. На севере (часть, соответствующая современной Евразии) и на юге (Австралия и Антарктида) климат был более влажными.







Наземная жизнь в триасовый период
В предшествующий пермский период преобладали амфибии, но в Триасе отмечался рост численности рептилий, в частности, архозавров и терапсид («звероящеров»). Точно не известно по какой причине архозавры доминировали над своими «звероподобными» родственниками терапсидами. Архозавры эволюционировали к среднему триасу в первых настоящих динозавров, таких как эораптор и герреразавр. Некоторые из них, однако, пошли в другом направлении, превратившись в первых птерозавров (эудиморфодон) и разнообразных примитивных крокодилов, некоторые из которых передвигались на двух ногах и питались растительностью. Терапсиды тем временем постепенно уменьшались в размерах. Первые млекопитающие позднего триасового периода (эозостродон и синоконодон) представляли из себя маленьких существ величиной с мышь.
Герреразавр — один из первых динозавров
Морская жизнь в Триасе
Поскольку пермское вымирание опустошило океаны, триасовый период был благоприятен для развития ранних морских рептилий. К ним относятся не только не классифицируемые, одновидовые роды, такие как плакодус и нотозавр, но и многочисленные плезиозавры и ихтиозавры. Некоторые ихтиозавры достигли огромных размеров, например шонизавр был 15 метров в длину и весил около 30 тонн! Огромный океан Панталасса постепенно заселялся новыми видами рыб, а также примитивными существами, такими как кораллы и головоногие моллюски.
Растительность триасового периода
Саговник Румфа
В триасовом периоде растительность не была такой пышной и распространенной как в более поздние юрский и меловой периоды. Тем не менее наземная растительность активно развивалась. Доминирующими группами растений были: саговники, папоротники, гинкгоподобные деревья и голосеменные растения. Одна из причин, по которой в этот период не было травоядных крупных размеров (как например более поздний брахиозавр), заключается в том, что растительности было не достаточно, чтобы их прокормить.
Триасово-юрское вымирание
Не самое известное массовое вымирание между триасовым и юрским периодами было не таким значительным как более раннее пермско-триасовое и более позднее мел-третичное вымирание. Тем не менее исчезли различные роды морских рептилий, амфибий и некоторые ветви архозавров. Достоверно не известно что послужило причиной этого вымирания, возможно, это была возросшая вулканическая активность, глобальное похолодание климата, падение метеоритов или какое-то сочетание этих факторов.
Почему динозавры так называются? (2 фото)
Почему динозавров так назвали
Для мировой науки динозавры были открыты в начале XIX века. В 1815 году в британском графстве Оксфордшир были обнаружены фрагмент нижней челюсти неизвестного на тот момент животного, а также несколько костей. Геолог Уильям Баклэнд назвал это существо мегалозавром, то есть огромным ящером. Несколько лет спустя, он доложил о своей находке в Королевском геологическом обществе. Через два года после его выступления хирург из Льюиса Гидеон Мэнтл представил этому же сообществу зубы еще одного неизвестного ископаемого животного, которые были найдены в графстве Суссекс. Этот странный житель был назван игуанодоном – за сходство находки с зубом ящерицы игуаны. А чуть позднее Мэнтл рассказал о новом существе – гилеозавре. А вот название динозавр появилось в 1842 году с подачи британского палеонтолога Ричарда Оуэна. Именно он сделал вывод о сходстве между найденными ранее фрагментами останков и отличии их строения от современных животных. Ученый понял, что это не рептилии и что они не принадлежат к классу современных пресмыкающихся
Анализируя находки, англичанин также принял во внимание гигантский размер существ. В итоге он назвал их динозаврами — этот термин происходит от двух древнегреческих слов: δεινός — «страшный», «ужасный», «странный» и σαύρα — «ящер», «ящерица»
Получается, динозавр – это страшный и странный ящер. Продолжив изучение вымерших животных, Оуэн привлек к доисторическим обитателям внимание политиков и ученых. Благодаря ему палеонтология получила дополнительное государственное финансирование. За свою жизнь Оуэн выпустил порядка 20 научных трудов, часть из которых была издана в виде большого сборника «История о древних рептилиях Британии».
Как называются различные виды динозавров
Рассмотрим названия различных видов динозавров: на основании чего они были придуманы. К примеру, анкилозавр происходит от древнегреческих ἀγκύλος — «согнутый» и σαῦρος — «ящер». По своему строению он действительно напоминал огромное согнутое существо. Слово «апатозавр» произошло от слияния ἀπάτη — «обман» и σαυρος — «ящер». Это связано с тем, что изучавшие его ученые изначально дали динозавру не то название, приписав ему чужую голову. «Обманчивый ящер» — подходящее прозвище для существа, которое ассоциируется с одной из самых больших ошибок в сфере изучения динозавров. Велоцираптор означает «быстрый охотник». Название происходит от латинских velox и raptor. Представители этого рода хищных двуногих динозавров были прирожденными бегунами. Животные, на которых они нападали, почти не имели шансов на спасение. Интересное происхождение названия у галлимима. Это слово появилось на стыке латинских gallus — «петух» и mimus — «имитатор». Эти существа имели беззубый клюв, длинные тонкие ноги и шею, изогнутую в форме буквы S. Возможно, такое строение напомнило ученым петуха. Название зухомим является союзом двух латинских слов: such — «крокодил» и mimus — «имитатор». Этот огромный рыбоядный динозавр отличался длинной, узкой и низкой мордой со множеством зубов, расположенных на конце челюсти – прямо как у крокодила. С тираннозавром все более-менее ясно: название произошло от древнегреческих τύραννος — «тиран» и σαῦρος — «ящер». Это существо было одним из самых крупных сухопутных хищных ящеров из тех, что когда-либо жили на нашей планете. По своим повадкам он был настоящим тираном, угнетателем и деспотом. А вот эдмонтозавр получил название в честь города Эдмонтона, так как его кости были обнаружены на территории южной Канады. Это животное считается одним из крупнейших гадрозавров: его рост доходил до 13 метров, а вес – до 4 тонн.